Dieser Prozess involviert lichtempfindliche Zellen (Fotorezeptoren) in der Retina, einer dünnen Schicht innerhalb des Auges. Die Rezeptoren erfassen Photonen und wandeln ihre Energie in Signale um, die vom Auge in den okzipitalen (visuellen) Kortex transportiert werden. Der visuelle Kortex ist der Bereich des Gehirns, der diese Signale als Sehen interpretiert.
Die Farben, die wir sehen, entstehen nicht aus einer Mischung aus Rot, Grün und Blau wie bei einem Monitor, sondern durch Vergleiche in einer nachgeordneten Ebene des Farbsehens.
Farbsehen spielt sich auf zwei Ebenen ab:
Als ob diese Maschinerie nicht schon komplex genug klingt: Das menschliche Farbsehen wird als trichromatisch bezeichnet. Wir besitzen drei unterschiedliche Klassen von Fotorezeptor-Zellen – Zapfen – die in einem unregelmäßigen Mosaik auf der Rückseite der Retina angeordnet sind.

Jede Zapfenklasse enthält ein Pigment, das maximal empfindlich gegenüber einem bestimmten Bereich des sichtbaren Spektrums ist:

Werfen wir einen Blick auf das Farbsehen der Dichromaten – z.B. eines Hundes. Dichromaten habe zwei Zapfen-Pigmente, eines für kurzwelliges bis UV-Licht (S) und eines für Lichtwellen im mittleren Bereich (M).
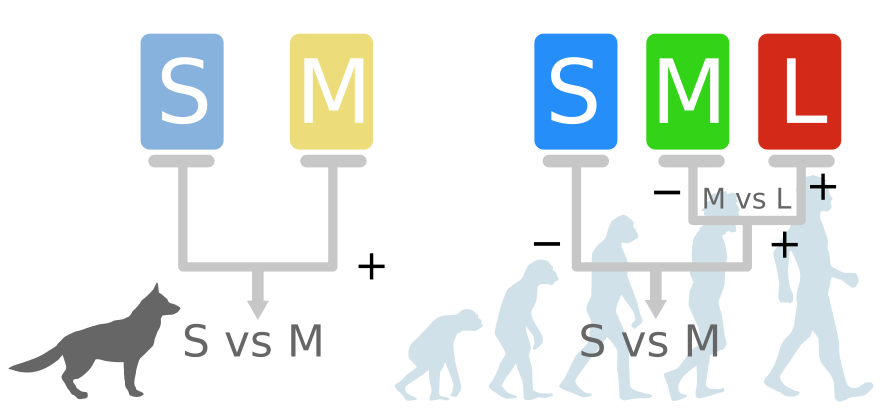
Durch einen komplexen Prozess, der durch die Sammelleidenschaft der Zapfen angestoßen wird, entstehen die Mischfarben in unserem Kopf: In einer einfachen Schaltung wird S mit M verglichen. Kommen mehr Photonen auf dem M-Zapfen an, sieht der Hund Gelb, ist das Signal stärker auf dem S-Zapfen, sieht der Hund Blau. In dem Bereich um 480 nm sieht der Hund kein Grün und keine Mischung, sondern Abstufungen von Grau.
Das Ergebnis des Vergleichs zwischen kurzen und langen Wellenlängen wird als Ausgabe an die Ganglien-Zellen geleitet.
Das ist die grundlegende Schaltung – auch in der Retina der Primaten.
Zunächst einmal funktioniert das Farbsehen beim Menschen nicht anders als beim Hund – obwohl wir Trichromaten sind.
Irgendwann in der Evolution der Primaten kam es zu einem Split des Pigments im M-Zapfen, der zu zwei neuen Seh-Pigmenten führte: Eines für das mittelwellige Licht um 530 nm und eines für das etwas langwelligere Licht so um 560 nm.
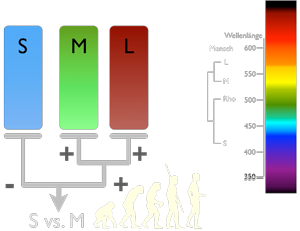
Das verbessert zwar die Analyse der mittleren Wellenlängen, aber die Schaltung macht keinen Unterschied zwischen M- und L-Zäpfchen. L und M werden gleichbehandelt und in einer erster Ebene einfach addiert. Am Ende ist die Ausgabe dieselbe wie beim Hund: Ein Vergleich zwischen kurzen und mittleren/längeren Wellenlängen. Diese Verrechnung erfolgt schon bald hinter den Rezeptoren, noch in der Augennetzhaut (Retina)
Dann kommt allerdings noch ein zweiter 1 Schaltkreis ins Spiel. Hier werden die Signale auf M gegen die Signale auf L verglichen. S wird nicht mit M oder L verglichen. Hier ist die Forschung dem Prozess des Farbsehens aber noch nicht ganz auf die Schliche gekommen.
Unter den Frauen findet man eine verschwindend kleine Anzahl von Tetrachromaten. Tetrachromaten haben nicht drei, sondern vier verschiedene Zapfentypen, wobei der vierte Typ irgendwo zischen Rot und Grün im Orangen Bereich zu liegen scheint.
Die Pigmente der roten und grünen Zapfen liegen im X-Chromosom, von dem bei Frauen zwei zu finden sind. So entsteht die Möglichkeit, dass sich die Zapfen unterschiedlich entwickeln. Bei den meisten Frauen allerdings liegen die beiden Zapfen aber so nah zusammen, dass keine besondere Farbwahrnehmung entsteht.
Mithilfe eines genetischen Scans können wir zwar herausfinden, ob eine Frau über eine vierte Zäpfchenart verfügt – aber es ist schwer nachzuweisen, dass sie über ein herausragendes Farbempfinden verfügt.
Etwa 8% der weißen europäischen/amerikanischen Männer hat eine Rot-Grün-Schwäche. Die meisten dieser Männer haben neben dem blauen Standardzapfen entweder zwei rote oder zwei grüne Zapfen.
Jay Neitz color vision researcher at the Medical College of Wisconsin.
